
编注:相信不少人对“舌尖、舌侧、舌根对应不同风味的感知”这一理论有印象,然而这个理论是真的吗?本文将梳理味觉运作的机理,破除一些关于味觉的错误认识。
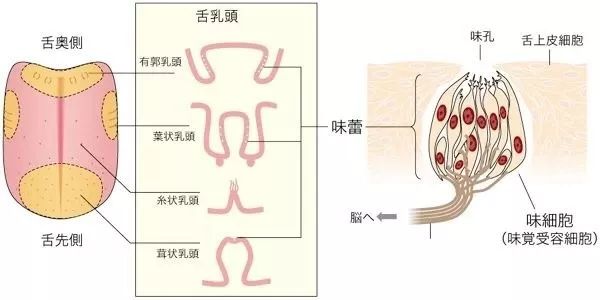
图片来自旦部幸博对咖啡的相关研究报告在舌头的上表面,有许多小的突起,称为舌乳头。味蕾实际上是感知味觉的器官,聚集在舌乳头中。人类的味觉感受器是覆盖在舌面上的味蕾。味蕾主要分布在舌头表面、上颚的黏膜中和喉咙周围,通常由大概50个细胞构成,其中包括一部分成熟味觉细胞与尚未成熟的味觉细胞,还有支持细胞与传导细胞等。
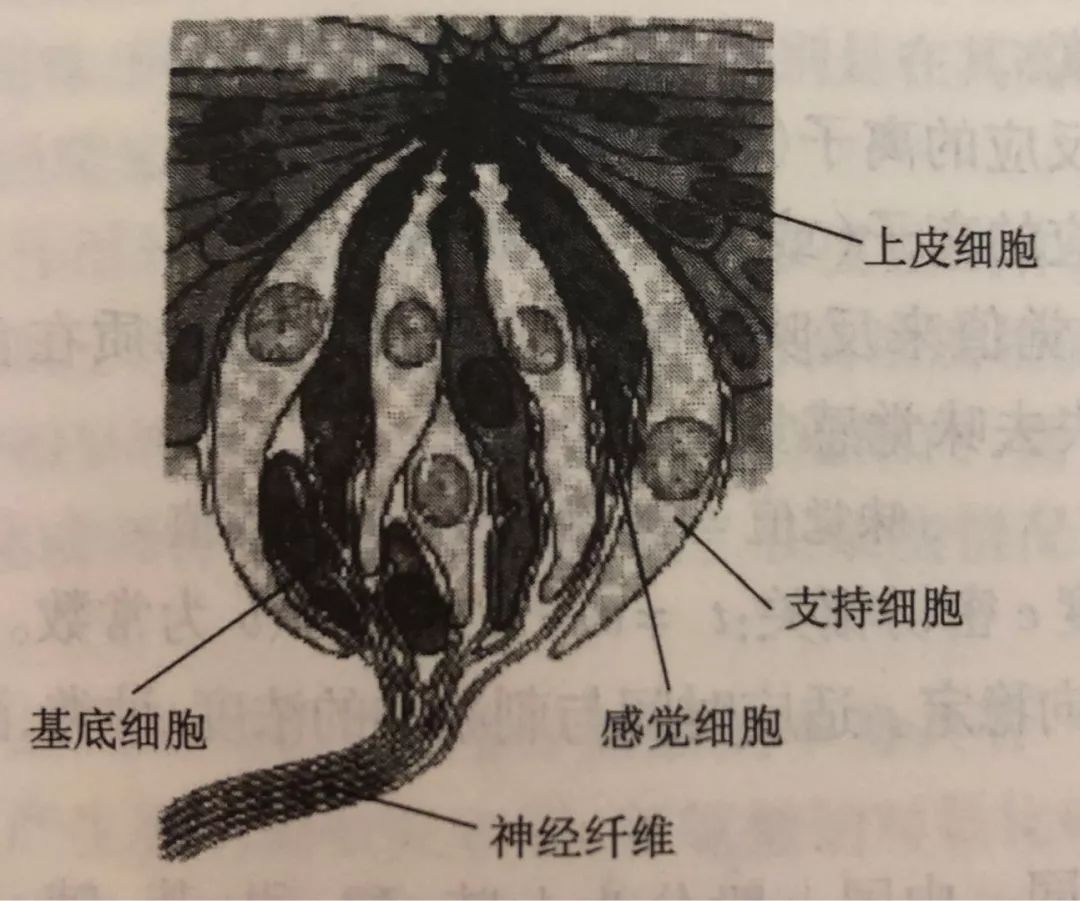
图片来自高等学校专业教材《食品感官分析》而我们的味蕾则分布在不同的味觉乳头上,这些味蕾由于乳头结构不同,导致对味觉的感知的敏感度不同,但这与流传尚久的“味觉地图”相违背,实际上每一个味蕾都可以感知到各种风味物质。只不过敏感度不同,且各类不同乳头在舌面分布的密集程度也有不同。舌乳头有四种类型:轮廓乳头形状像被护城河围绕的城堡般,被凹槽包围,仅舌根周围约有10个。在乳突周缘?侧上存在大量的味蕾,每个突起有数百到千个,并且利用在凹槽中积聚的液体(分泌液,例如唾液和食物来源的液体)感知溶解的味道物质。凹槽)中有一个分泌腺,称为埃伯纳腺,埃伯纳腺的分泌物将凹槽中的残留物质冲洗掉,从而防止了味物质永久保留。叶状乳头百褶的舌乳头,呈褶状,仅出现在舌根处,靠近根部。叶状乳头在突起的侧面也有味蕾,但数量远少于乳突周缘,每个突起约十几个。埃伯纳腺也存在于叶状乳头褶皱的底部,其作用也是分泌液将残留物质冲洗掉。丝状乳头舌头乳头,角质较薄,当用肉眼观察时是白色的凸起物。存在于整个舌头的上表面。丝状乳突中没有味蕾,它们不参与基本的味觉感知。丝状乳头是舌头的真正“粒状”性质,在人们舔食物时,丝状乳头发挥了类似于锉刀的作用。菌状乳头它类似于丝状乳头,但未角质化,血管可用肉眼直接观察到,尖端呈红色。像丝状乳头一样,它存在于整个舌头的上表面,但特别集中在前表面。真菌状乳突的尖端通常具有一个或几个味蕾,但其中一些缺失且没有味蕾。舌乳头分布的这些差异导致舌上味蕾分布的偏差。在舌头中,靠近根部(约2,200个,多分布在外接乳头上),在舌头后部(约1,300个,多分布在叶状乳头上),在舌头上表面(约1,100个,多分布在蘑菇状乳头上)由于存在味蕾,这些部分大大有助于味觉。除舌头外,口腔中还存在约2300个味蕾,据说它们与味觉有关。在舌头的表面上,舌头上皮细胞折叠形成上皮组织,但味蕾缺少该上皮层,并有称为味孔的孔。在这个味孔下,几个味觉细胞聚集形成一个味蕾。每个味觉细胞都与味觉神经相连,当味觉物质作用于味觉细胞顶部的微绒毛时,信号会通过味觉神经发送至大脑,从而识别味觉。每个味觉神经所属的神经系统取决于味蕾的位置。带有蘑菇状乳突的舌头的前三分之一是面神经(第七对颅神经),具有叶状乳头和轮廓乳头的舌头的后三分之二是舌咽神经(第九对颅神经),会厌和咽大部分的喉咙受迷走神经(第十对颅神经)控制。这些神经连接到称为孤束核的延髓部分,这些部分将信息传递到大脑。舌咽神经在味觉感知中起着最大的作用。味觉的强度和出现味觉的时长,与物质的水溶性密切相关,实际上绝对不溶于水的物质是没有味道的。只有可溶于水的物质才能刺激味觉神经。而水溶性好的物质味觉产生更快,但也更短暂,这也是某些咖啡的明亮酸质转瞬即逝的来源,也是谷物甜较为短暂的原因。而水溶性不好的物质,味觉产生更慢,也就是所谓的余韵与回甘。而风味的强度则与味觉物质的浓度与阀值密切相关。
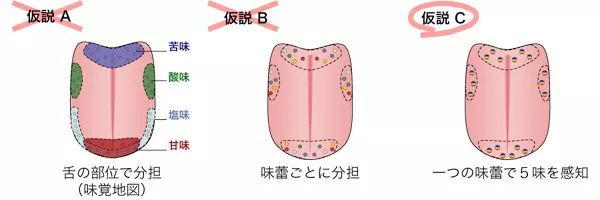
图片来自旦部幸博对咖啡的研究报告上图阐述了味觉相关研究的过程a,这是最常见的“味觉地图”b,这是后来流行的不同味蕾单一对风味的感知。c,正确的情况是每一个味蕾都可以感知到不同风味,只是不同区域的敏感度不一样。除去我们常见的基本味“酸、甜、苦、咸、鲜”外,实际上,辣味说是触觉其实并不严谨,辣味是味蕾对辣味的一种感知,它与味蕾对温度的感知是共用同一个味觉神经,所以辣味同时会给我们一种灼烧的感觉。而涩感则由物质对口腔黏膜蛋白质的变性而产生的收敛感引发。那么,“一个味蕾”如何对应基本味?一个味蕾中存在多个味觉细胞,并通过味觉神经向大脑发送信号,一个味觉细胞专门对应一种基本味觉的接收,并从那里连接到单一特定于味觉的味觉神经,并将其作为“一种味觉”传递到大脑,不同的味道使用了不同的味觉神经。在味觉细胞的顶部,有一种蛋白质(味觉受体)与细胞表面(细胞膜上)的特定味觉成分结合。被动物和人类感知为味道的化学物质称为呈味物质。对于每种基本风味,都有一种产生该风味的呈味物质,但是某些呈味物质会触发多种味觉受体,例如某些糖精既引起苦味又引起甜味。另外,呈味物质需要达到一定的浓度才能被感知,如果浓度低于我们的基础味阈,则不能被感知。可以感知到味道的浓度称为阈值,阈值越小,味道物质的作用越强。当味道物质包含有多重味道的情况下,每种味道的阈值是不同的。例如,上述糖精的甜度阈值小于苦味,因此低浓度的糖精主要感觉到甜味,而高浓度则会感到苦味的增加。引起甜味的甜味物质包括蔗糖和葡萄糖等糖,阿斯巴甜和糖精等人造甜味剂以及中草药甘草中所含的甘草甜素等糖苷。一种称为甜蛋白的蛋白质也具有甜味,例如莫内林和仙茅素都属于甜蛋白。在某些不常见的情况下,某些无机金属化合物(如醋酸铅)(有毒)和某些有机溶剂(如氯仿CHCl3)也具有甜味。在糖类中,聚合度低且分子量较小的单糖例如葡萄糖和果糖,以及二糖例如蔗糖和麦芽糖,但是不产生甜味的例如淀粉,这种高聚合度的多糖则失去甜味。分子生物学研究的结果,在遗传水平上已经鉴定出甜味,鲜味和苦味受体,并且已经鉴定出酸性受体的候选分子。咸受体仍是未知的。以下专业内容来自于旦部幸博:甜味,鲜味和苦味的受体都是被称为“七跨膜蛋白”的膜蛋白,具有七个穿透细胞膜的区域。它属于G蛋白偶联受体(GPCR)受体组,因为它与称为三聚体GTP结合蛋白的蛋白相连。这些甜味,鲜味和苦味受体蛋白可大致分为两种类型:具有大的细胞外区域的蛋白和不具有该区域的蛋白,而前者则分为T1R家族(或TAS1R,味觉受体类型)后者称为T2R家族(TAS2R,味觉受体2型)。T1R家族进一步包括三种类型,T1R1至T1R3,其中T1R1和T1R3的组合构成鲜味受体,并且T1R2和T1R3的组合分别构成甜味受体。我明白 T2R家族成员更多,包括人类26个和小鼠35个。其中,分析了多个T2R家族基因(例如T2R5)在味觉接收中的功能,并且证明了T2R家族起苦味受体的作用。T2R家族在作为苦味受体时是否需要结合多个T2R分子尚不清楚,但与甜味和鲜味一样,它可能不会以任何方式形成复合物。一方面,酸味和咸味的受体被认为是离子通道型分子,是一种与温度和辣味等受体有关的,被称为TRP通道(瞬态受体电位通道)的通道型分子的同类分子。一般来说TRP通道型分子是六跨膜膜蛋白,它并非是单一类型的通道分子,而是多种类型的通道分子共同形成的复合物,被认为是一个通道分子。虽然还不能确认酸味与咸味的受体有关的分子,我们猜想酸味受体和咸味受体分别依靠氢离子和钠离子的浓度梯度从细胞外向细胞内移动,形成类似的离子通道各自进行作用。实际上最近也有关于酸味受体的候选物的报告。在小鼠中,有迄今为止一直被认为是TRP通道分子的PKD2L1,与被称为PKD1L3的拥有大胞外区十一次贯通膜蛋白所形成的复合物。有猜想认为它们是通道型的味觉受体中酸味受体,它们也被认为是哺乳动物的酸味受体的有力候补物之一。下表显示了迄今为止确定的典型味觉受体与结合它们的典型味觉物质(配体)之间的对应关系。许多苦味物质的名称可能不为人所熟悉,但是由于它们都被发现具有强烈的苦味,因此被用于苦味研究目的,或为避免误被加入服杀虫剂和其他物质。例如苯甲地那铵,在浓度只有0.01ppm的情况下已经能被感知到苦味,已经被吉尼斯世界纪录认证为世界上最苦的物质。一些苦味物质的受体未知,将在后面的部分中进行解释。味觉受体及其配体

-甜味-受体:T1R2 + T1R3配体:糖(蔗糖,葡萄糖,果糖,麦芽糖),人造甜味剂(糖精,阿斯巴甜,乙酰磺胺酸钾,甜蜜素),甜蛋白(莫内林,仙茅素)-鲜味 -受体:T1R1 + T1R3配体:氨基酸(L-谷氨酸),核酸(肌苷)-苦味-受体:T2R5配体:环己酰亚胺受体:T2R4、8、44配体:苯甲地那铵受体:T2R16配体:水杨素受体:T2R38配体:苯基硫脲(PTC)受体:T2R43、44配体:糖精受体:不明配体:奎宁,士的宁,阿托品,咖啡因- 酸味 -受体:pkd2l1 + pkd1l3(?)配体:酸(柠檬酸,酒石酸,乙酸)(编注:2019年9月29日,生命科学期刊Cell发表了一篇名为《Sour Sensing from the To
ngue to the Brain》的研究报告,指出酸味受体为离子通道OTOP-1)

参考文献《化学总说14:味道与气味的化学》(日本化学会1976)稻田仁、富永真琴(2007)细胞工学26,890-893旦部幸博《咖啡的科学》旦部幸博个人博客周家春《食品感官分析》(中国轻工业出版社)

-欢迎关注作者公众号 -▼

版权信息:以上文章内容转载自【有脾气的烘焙师】,转载请联系原作者。作者:Roaster北巷侵权请联系:weixin@coffeesalon.com投稿合作:tips@coffeesalon.com

 手机浏览中华饮品届
手机浏览中华饮品届













